







People (including virtually all physicians) are constantly confused what cellular inflammation is. So I decided to take the opportunity to explain the concept in more detail.
There are two types of inflammation. The first type is classical inflammation, which generates the inflammatory response we associate with pain such as, heat, redness, swelling, pain, and eventually loss of organ function. The other type is cellular inflammation, which is below the perception of pain. Cellular inflammation is the initiating cause of chronic disease because it disrupts hormonal signaling networks throughout the body.
Definition of Cellular Inflammation
The definition of cellular inflammation is increased activity of the gene transcription factor know as Nuclear Factor-kappaB (NF-κB). This is the gene transcription factor found in every cell, and it activates the inflammatory response of the innate immune system. Although the innate immune system is the most primitive part of our immune response, it has been resistant to study without recent breakthroughs in molecular biology. In fact, the 2011 Nobel Prize in Medicine was awarded for the earliest studies on the innate immune system and its implications in the development of chronic disease.
There are several extracellular events through which NF-κB can be activated by distinct mechanisms. These include microbial invasion recognized by toll-like receptors (TLR), generation of reactive oxygen species (ROS), cellular generation of inflammatory eicosanoids, and interaction with inflammatory cytokines via defined cell surface receptors. We also know that several of these initiating events are modulated by dietary factors. This also means that appropriate use of the diet can either turn on or turn off the activation of NF-κB. This new knowledge is the foundation of anti-inflammatory nutrition (1-3).
Understanding Cellular Inflammation
Although the innate immune system is exceptionally complex, it can be illustrated in a relatively simple diagram as shown below in Figure 1.
Figure 1. Simplified View of the Innate Immune System
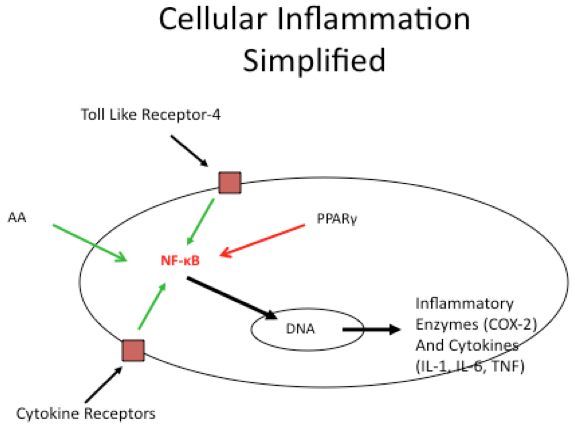
Essential fatty acids are the most powerful modulators of NF-κB. In particular, the omega-6 fatty acid arachidonic acid (AA) activates NF-κB, whereas the omega-3 fatty acid eicosapentaenoic acid (EPA) does not (4). Recent work suggests that a subgroup of eicosanoids known as leukotrienes that are derived from AA may play a significant factor in NF-κB activation (5,6)
Extracellular inflammatory cytokines can also activate NF-κB by their interaction with specific receptors on the cell surface. The primary cytokine that activates NF-κB is tumor necrosis factor (TNF) (7). Toll-like receptors (TLR) are another starting point for the activation of NF-κB. In particular, TLR-4 is sensitive to dietary saturated fatty acids (8). The binding of saturated fatty acids to TLR-4 can be inhibited by omega-3 fatty acids such as EPA. Finally ROS either induced by ionizing radiation or by excess free radical formation are additional activators of NF-κB (9).
Anti-inflammatory Nutrition To Inhibit Cellular Inflammation
Anti-inflammatory nutrition is based on the ability of certain nutrients to reduce the activation of NF-κB.
The most effective way to lower the activation of NF-κB is to reduce the levels of AA in the target cell membrane thus reducing the formation of leukotrienes that can activate NF-κB. Having the patient follow an anti-inflammatory diet, such as the Zone Diet coupled with the simultaneous lowering omega-6 fatty acid intake are the primary dietary strategies to accomplish this goal (1-3).
Another effective dietary approach (and often easier for the patient to comply with) is the dietary supplementation with adequate levels of high-dose fish oil rich in omega-3 fatty acids, such as EPA and DHA. These omega-3 fatty acids taken at high enough levels will lower AA levels and increase EPA levels. This change of the AA/EPA ratio in the cell membrane will reduce the likelihood of the formation of inflammatory leukotrienes that can activate NF-κB. This is because leukotrienes derived from AA are pro-inflammatory, whereas those from EPA are non-inflammatory. The increased intake of omega-3 fatty acids is also a dietary approach that can activate the anti-inflammatory gene transcription factor PPAR-γ (10-12), decrease the formation of ROS (13) and decrease the binding of saturated fatty acids to TLR-4 (14). This illustrates the multi-functional roles that omega-3 fatty acids have in controlling cellular inflammation.
A third dietary approach is the adequate intake of dietary polyphenols. These are compounds that give fruits and vegetables their color. At high levels they are powerful anti-oxidants to reduce the generation of ROS (15). They can also inhibit the activation of NF-κB (16).
Finally, the least effective dietary strategy (but still useful) is the reduction of dietary saturated fat intake. This is because saturated fatty acids will cause the activation of the TLR-4 receptor in the cell membrane (8,14).
Obviously, the greater the number of these dietary strategies implemented by the patient, the greater the overall effect on reducing cellular inflammation.
Clinical Measurement of Cellular Inflammation
Since cellular inflammation is confined to the cell itself, there are few blood markers that can be used to directly measure the levels of systemic cellular inflammation in a cell. However, the AA/EPA ratio in the blood appears to be a precise and reproducible marker of the levels of the same ratio of these essential fatty acids in the cell membrane.
As described above, the leukotrienes derived from AA are powerful modulators of NF-κB. Thus a reduction in the AA/EPA ratio in the target cell membrane will lead to a reduced activation of NF-κB by decreased formation of inflammatory leukotrienes. The cell membrane is constantly being supplied by AA and EPA from the blood. Therefore the AA/EPA ratio in the blood becomes an excellent marker of the same ratio in the cell membrane (17). Currently the best and most reproducible marker of cellular inflammation is the AA/EPA ratio in the blood as it represents an upstream control point for the control of NF-κB activation.
The most commonly used diagnostic marker of inflammation is C-reactive protein (CRP). Unlike the AA/EPA ratio, CRP is a very distant downstream marker of past NF-κB activation. This is because one of inflammatory mediators expressed in the target cell is IL-6. It must eventually reach a high enough level in the blood to eventually interact with the liver or the fat cells to produce CRP. This makes CRP a more long-lived marker in the blood stream compared to the primary inflammatory gene products (IL-1, IL-6, TNF, and COX-2) released after the activation of NF-κB. As a consequence, CRP is easier to measure than the most immediate inflammatory products generated by NF-κB activation. However, easier doesn’t necessarily translate into better. In fact, an increase AA/EPA ratio in the target cell membrane often precedes any increase of C-reactive protein by several years. An elevated AA/EPA ratio indicates that NF-κB is at the tipping point and the cell is primed for increased genetic expression of a wide variety of inflammatory mediators. The measurement of CRP indicates that NF-κB has been activated for a considerable period of time and that cellular inflammation is now causing systemic damage.
In Summary
I believe the future of medicine lies in the control of cellular inflammation. This is most effectively accomplished by the constant application of anti-inflammatory nutrition. The success of such dietary interventions can be measured clinically by the reduction of the AA/EPA ratio in the blood.

References:
- Sears B. The Anti-Inflammation Zone. Regan Books. New York, NY (2005).
- Sears B. Toxic Fat. Thomas Nelson. Nashville, TN (2008).
- Sears B and Riccordi C. “Anti-inflammatory nutrition as a pharmacological approach to treat obesity.” J Obesity doi:10.1155/2011/431985 (2011).
- Camandola S, Leonarduzzi G,Musso T, Varesio L, Carini R, Scavazza A, Chiarpotto E, Baeuerle PA, and Poli G. “Nuclear factor kB is activated by arachidonic acid but not by eicosapentaenoic acid.” Biochem Biophys Res Commun 229:643-647 (1996).
- Sears DD, Miles PD, Chapman J, Ofrecio JM, Almazan F, Thapar D, and Miller YI. “12/15-lipoxygenase is required for the early onset of high fat diet-induced adipose tissue inflammation and insulin resistance in mice.” PLoS One 4:e7250 (2009).
- Chakrabarti SK, Cole BK, Wen Y, Keller SR, and Nadler JL. “12/15-lipoxygenase products induce inflammation and impair insulin signaling in 3T3-L1 adipocytes.” Obesity 17:1657-1663 (2009).
- Min JK, Kim YM, Kim SW, Kwon MC, Kong YY, Hwang IK, Won MH, Rho J, and Kwon YG. “TNF-related activation-induced cytokine enhances leukocyte adhesiveness: induction of ICAM-1 and VCAM-1 via TNF receptor-associated factor and protein kinase C-dependent NF-kappaB activation in endothelial cells.” J Immunol 175: 531-540 (2005).
- Kim JJ and Sears DD. “TLR4 and Insulin Resistance.” Gastroenterol Res Pract doi:10./2010/212563 (2010).
- Bubici C, Papa S, Dean K, and Franzoso G. “Mutual cross-talk between reactive oxygen species and nuclear factor-kappa B: molecular basis and biological significance.” Oncogene 25: 6731-6748 (2006).
- Li H, Ruan XZ, Powis SH, Fernando R, Mon WY, Wheeler DC, Moorhead JF, and Varghese Z. “EPA and DHA reduce LPS-induced inflammation responses in HK-2 cells: Evidence for a PPAR-gamma-dependent mechanism.” Kidney Int 67: 867-874 (2005).
- Kawashima A, Harada T, Imada K, Yano T, and Mizuguchi K. “Eicosapentaenoic acid inhibits interleukin-6 production in interleukin-1beta-stimulated C6 glioma cells through peroxisome proliferator-activated receptor-gamma.” Prostaglandins LeukotEssent Fatty Acids 79: 59-65 (2008).
- Chambrier C, Bastard JP, Rieusset J, Chevillotte E, Bonnefont-Rousselot D, Therond P, Hainque B, Riou JP, Laville M, and Vidal H. “Eicosapentaenoic acid induces mRNA expression of peroxisome proliferator-activated receptor gamma.” Obes Res 10: 518-525 (2002).
- Mas E, Woodman RJ, Burke V, Puddey IB, Beilin LJ, Durand T, and Mori TA. “The omega-3 fatty acids EPA and DHA decrease plasma F(2)-isoprostanes.” Free Radic Res 44: 983-990 (2010).
- Lee JY, Plakidas A, Lee WH, Heikkinen A, Chanmugam P, Bray G, and Hwang DH. “Differential modulation of Toll-like receptors by fatty acids: preferential inhibition by n-3 polyunsaturated fatty acids.” J Lipid Res 44: 479-486 (2003).
- Crispo JA, Ansell DR, Piche M, Eibl JK, Khaper N, Ross GM, and Tai TC. “Protective effects of polyphenolic compounds on oxidative stress-induced cytotoxicity in PC12 cells.” Can J Physiol Pharmacol 88: 429-438 (2010).
- Romier B, Van De Walle J, During A, Larondelle Y, and Schneider YJ. “Modulation of signaling nuclear factor-kappaB activation pathway by polyphenols in human intestinal Caco-2 cells.” Br J Nutr 100: 542-551 (2008).
- Yee LD, Lester JL, Cole RM, Richardson JR, Hsu JC, Li Y, Lehman A, Belury MA, and Clinton SK. “Omega-3 fatty acid supplements in women at high risk of breast cancer have dose-dependent effects on breast adipose tissue fatty acid composition.” Am J Clin Nutr 91: 1185-1194 (2010).

Let Us Know What You Thought about this Post.
Put your Comment Below.